Major histocompatibility complex (MHC) molecules are tightly linked cluster of genes whose products are associated with intercellular recognition and self-non-self-discrimination. MHCs are present in every mammalian species. The MHC is referred to as the HLA complex in humans and the H-2 complex in mice.
Table of Contents
MHC Genes and Functions:
It is a collection of genes within a long stretch of DNA on chromosome 6, which codes for three classes of molecules
- Class I MHC genes: encode glycoproteins expressed on the surface of nearly all nucleated cells; the major function of the class I gene product is presentation of peptide antigens to TC cells.
- Class II MHC genes encode glycoproteins expressed primarily on APCs, where they present processed antigenic peptides to TH cells.
- Class III MHC genes encode various secreted proteins that have immune functions, including components of the complement system and molecules involved in inflammation (e.g. TNF, Heat Shock proteins).
MHC I
MHC Class I is found on all nucleated cells. MHC Class I mediates immune responses against endogenous antigens. Usually, these cells expressing MHC class I are viral-infected or tumor cells.
MHC Class I presents peptides that are 8 – 10 amino acids in size, which will then be recognized by the cytotoxic T cells.

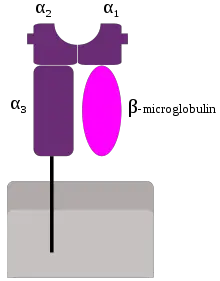
Structure of MHC Class I
- a chain anchored in plasma membrane by its hydrophobic transmembrane segment & hydrophilic cytoplasmic tail
- a chain has 3 external domains
- Homology between a3 & b2 microglobulin & constant regions in immunoglobulins
- Peptide-binding cleft
Class I MHC-Peptide Interaction and Presentation
Single nucleated cell expresses 105 copies of each class I molecule. Class I MHC will express many different peptides simultaneously on the surface. Each type of Class I (A, B, C in humans) bind a unique set of peptides & presents these peptides to CD8+ T-cells.
Endogenous Processing Pathway
- Class I MHC molecules bind peptides derived from endogenous antigens processed in the cytoplasm.
- Proteins enter the cytoplasm of cells either from phagocytosed microbes or from endogenous synthesis by microbes, such as viruses that reside in the cytoplasm of infected cells.
- Cytoplasmic proteins are unfolded, ubiquitinated, and degraded by proteasomes.
- Intracellular protein will be broken down into smaller peptides by a protease.
- The peptides will be transported into the endoplasmic reticulum by the TAP transporter, where the peptide bind with newly synthesized class I MHC molecules.
- The 8 – 10 amino acid residues will bind with MHC class I.
- The MHC class-I-antigen complex will migrate towards the cell surface and are presented to CD8+ T cells.
- CD8+ T cells may recognize the MHC-I Antigen complex and further processing takes place.

MHC II
The MHC class II protein is found on Antigen Presenting Cells (APCs). Class II MHC can bind to various peptides and presents these to CD4+ T cells. MHC class II mediates immune responses against exogenous antigens.
MHC class II will bind with amino acid residues that are 13 – 18 in size and will be recognized by T helper cells. These cells are phagocytic and can engulf an extracellular antigen.

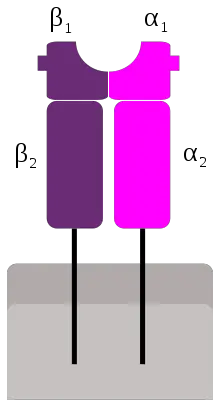
MHC II Structure
- It contains 2 different polypeptide chains: a chain (33 kDa) & a b chain (28 kDa)
- Each chain has 2 external domains
- Antigen binding cleft for processed antigens
- ab heterodimer “dimer of dimers.”
Class II MHC-Peptide Interaction and Presentation
Class II MHC molecules bind peptides derived from exogenous antigens internalized by phagocytosis or endocytosis & processed within the endocytic pathway. APCs can internalize antigens by phagocytosis &/or endocytosis. Macrophages do both; B cells use receptor-mediated endocytosis.
Exogenous processing pathway

- Microbial proteins enter intracellular vesicles called endosomes or phagosomes, which may fuse with lysosomes. The internalized antigen is degraded into peptides.
- APCs synthesize class II MHC molecules in the endoplasmic reticulum (ER).
- Each newly synthesized MHC-II carries an invariant chain or CLIP that binds to the peptide binding cleft, thus occupying the peptide binding region.
- This MHC Class II begins its transport to the cell surface in an exocytic vesicle, which then fuses with the endosomal vesicle containing broken-down peptides derived from ingested extracellular proteins.
- Endosomal vesicle contains a protein named DM, which removes CLIP. Thus, the cleft is available to accept peptides.
- Internalized antigen takes 1-3 hours to travel the endocytic pathway & appear on the cell membrane in the form of peptide-class II MHC complexes.
- If the MHC II molecule does not find a suitable peptide to bind, the empty molecule is unstable and is degraded by proteases in the endosomes.
- MHC-II will migrate to the phagolysosome, which will bind to components of 13- 18 amino acids in size.
- Once bound, the MHC class II will migrate to the membrane to display the antigen. A helper T cell will recognize the complex and trigger the appropriate response.
Difference between MHC I and MHC II
The major difference between MHC Class I and MHC Class II (including their antigen processing and presentation pathway) is summarized in this table:
| MHC Class I | MHC Class II | |
| Structure | MHC class I molecules consist of one membrane-spanning α chain (heavy chain) produced by MHC genes, and one β chain (light chain or β2-microglobulin) produced by the β2-microglobulin gene.
 | MHC class II molecules consist of two membrane-spanning chains, α and β, of similar size and both produced by MHC genes.
 |
| Types of APCs | MHC I glycoproteins are present in all nucleated cells. | MHC II glycoproteins are only present on specialised antigen-presenting cells (APCs), including macrophages that engulf foreign particles such as bacteria, dendritic cells that present antigen to T cells, and B cells that produce antibodies. |
| Nature of Antigen Presentation | MHC class I glycoproteins present endogenous antigens that originate from the cytoplasm. | MHC II proteins present exogenous antigens that originate extracellularly from foreign bodies such as bacteria. |
| Size of peptide | MHC Class I present 8-10 amino acid peptides | MHC Class II presents 14-18 amino acid peptides. |
| Responsive T Cells | Present antigen to cytotoxic T cell lymphocytes (CD8+ T Cells); | Present antigen to helper T cell lymphocytes; (CD4+ T cells). |
| Co-receptor responsible | Binds with CD8 coreceptors molecules on cytotoxic T cells | Binds with CD4 co-receptors molecules on helper T cells |
| Sources of Protein Antigens | Cytosolic proteins (mostly synthesized in the cell, may enter cytosol from phagosomes) | Endosomal/lysosomal proteins (mostly internalized from extracellular environment) |
| Enzymes Responsible for peptide generation | Cytosolic proteasome | Endosomal and lysosomal proteases (e.g., cathepsins) |
| Site of peptide loading of MHC | Endoplasmic reticulum | Specialized vesicular compartment |
| Molecules involved in transport of peptides and loading of MHC molecules | Chaperones, TAP in ER | Chaperones in ER; invariant chain in ER, Golgi and MHC Class II compartment/Class II vesicle; DM |
| End Result | Presentation of foreign-intracellular antigens or altered self-antigens; targets cell for destruction | Presentation of foreign extracellular antigens; induces antibody production, and attracts immune cells to area of infection |
{kind=link}
{kind=link}
Roles of MHC
- Development of humoral and cell-mediated immune response
- Antigen recognition by T cells: T cells recognize antigen only when it is combined with MHC molecule
- T cell receptor (TCR) can only bind to peptides processed from proteins not whole proteins.
- Peptides which are bound to MHCs are presented to T cells
- The main two groups of MHC important in adaptive T cell responses are MHC-I and MHC-II. MHC-I presents peptide to CD8 T cells and MHC-II presents peptide to CD4 T cells.
- Determining whether transplanted tissue will be histocompatible or histoincompatible
MHC Polymorphism and Significance
MHC genes are highly polymorphic (presence of multiple alleles at a given genetic locus within a species). The polymorphism is so great that NO two individuals in the usual outbreed population have exactly the same MHC genes and molecule.
- These polymorphic residues determine which peptides are presented by which MHC molecules
- This evolution of MHC polymorphism ensures that a population will not succumb to a microbe that mutates its proteins because at least some individuals will be able to mount effective immune responses to the peptide antigens of new introduced or mutated microbes.
- MHC expressed by an individual does not change over time but differ significantly from those expressed by another individual of the same species
- The different allele combinations make up the identity of the MHC for that person
- There are lots and lots of alleles and thousands of combinations, which is why finding someone to whose HLA markers match up w/ another person is so difficult but it’s not impossible, there are thousands of transplants every year.
References and further reading:
Hlo sir…
Could you suggest me about antibiotic sensitivity test for bacteria…. I can’t found anything about this test
I wana find out the bacteria that produce antibiotics…so plz sir help me..
Hi, you can find antibiotic producing bacteria by the following procedure.
1. select source mean sample area.
2. make lawn of bacteria and spread very little amount of sample over lawan.
3. incubate for 24hrs and then check result.
4. there will lot of growth on the medium if you find zone of inhibition on plate, take that colony for sure that one is antibiotic producers.
Source of protein antigens: I thought that MHC1 presented peptides from internal proteins, the majority from failed manufactured proteins in the ER. MHC11 are for presenting peptides that have been internalised such as viruses . Haven’t you got them the wrong way round?
Dear Henry Mannings
Thank you so much for your comment and making me aware about the mistakes done while posting this blog. With your help, now i became able to correct the exchanged information. I will be looking for your feedback and suggestions in future too.
Please help me to answer these questions.
Why is always mycobacterium tuberculosis( acid fast bacilli) is not matching with gram staining ? And what category is standing for as bacteria gram activities ?
Hello sir ,
Why mycobacteria tuberclusosis does not staining with gram staining ?
Which gram staining is mycobacteria tuberclusosis ?
Very nice information sir, You have simplified immunology topic MHC class of membrane proteins in simplified way, very thankful to you
Hello sir, can you help me suggest a reason for why different MHC class 1 molecules between donor and recipient cells can lead to rejection of a transplanted organ or tissue ?
Nice information and your the best for microbiology topics.
Thanks for this helpful post
MHC explanation needs some more clarity , MHC 4th type is also there but in ur topic only 3 are there sir. Explain with common n simple examples.Tq sir.