Lac Operon: Mechanism and Regulation
Lac Operon: Mechanism and Regulation
The lac operon is a well-known example of an inducible gene network that regulates the transport and metabolism of lactose in Escherichia coli. It encodes the genes for the internalization of extracellular lactose and then its conversion to glucose.
The lactose operon of E. coli is turned ON only when lactose is available (and glucose, the preferred energy source, is absent). When there is an absence of lactose the transcription of the lac operon genes is blocked by a repressor protein (as there will be no use of operon’s gene products).
Structure of the lac operon
The lac operon consists of a promoter (P) and operator (O) region followed by three structural genes lacZ, lacY, and lacA in the downstream. A regulatory gene lacI (I) preceding the lac operon is responsible for producing a repressor (R) protein.
| Structural gene | Enzyme | Function |
|---|---|---|
| lacZ | β-galactosidase (B) | It transforms lactose into allolactose and also catalyzes the conversion of lactose to glucose and galactose. |
| lacY | permease (P) | Membrane channel protein required to uptake lactose from the environment |
| lacA | thiogalactoside transacetylase | It rids the cell of toxic thiogalactosides that also get transported by lacY. |
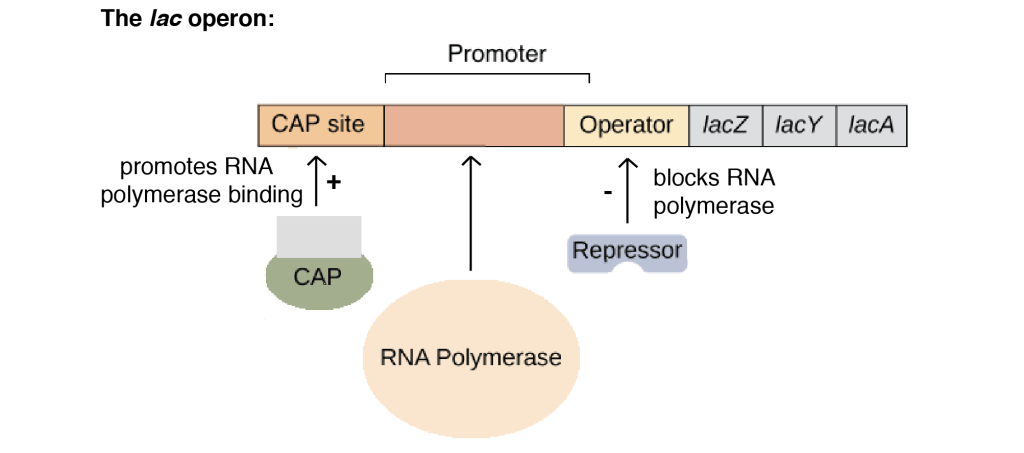 In addition to structural genes, the lac operon also contains a number of regulatory DNA sequences. These are regions of DNA to which particular regulatory proteins can bind, controlling transcription of the operon.
In addition to structural genes, the lac operon also contains a number of regulatory DNA sequences. These are regions of DNA to which particular regulatory proteins can bind, controlling transcription of the operon.
The promoter is the binding site for RNA polymerase, the enzyme that performs transcription. The lac promoter is located at 5′ end of lacZ and directs transcription of all the three genes as a single mRNA. This mRNA is translated to give three protein products (shown in the table below)
Structural gene | Enzyme | Function |
lacZ | β-galactosidase (B) | It transforms lactose into allolactose and also catalyzes the conversion of lactose to glucose and galactose. |
lacY | permease (P) | Membrane channel protein required to uptake lactose from the environment |
lacA | thiogalactoside transacetylase | It rids the cell of toxic thiogalactosides that also get transported by lacY. |
Prokaryotic genes expression is very often controlled by extracellular signals i.e. substrates present in the growth medium. Two regulatory proteins communicate these signals with the genes:
- Positive regulators (activators): Activator increases transcription of the regulated genes. In lac operon, activator ( called CAP) acts as a glucose sensor. It activates the transcription of the operon when glucose is absent/low.
- Negative regulators (repressor): Repressor decreases or eliminates transcription of genes. In lac operon, repressor acts as a lactose sensor. Lac repressor is encoded by the lacI gene. Lac repressor prevents transcription of structural genes for lactose metabolism when lactose is not available by tightly binding to the operator region.
DO YOU KNOW?
Jacques Monod, together with François Jacob has formulated lac operon model for the regulation of gene expression in the late 1950s. The two of them, together with their colleague André Lwoff were awarded with“ The Nobel Prize in Physiology or Medicine” in 1965. Since its discovery, lac operon has been serving as a model system for understanding different aspects of gene regulations.
Mechanism of the lac operon
When lactose is absent
The lac repressor senses the presence of lactose (more precisely allolactose-an isomer of lactose) in the medium.
In the absence of allolactose (A) the repressor protein (R) binds to the operator region (O) and blocks the RNA polymerase from transcribing the structural genes.
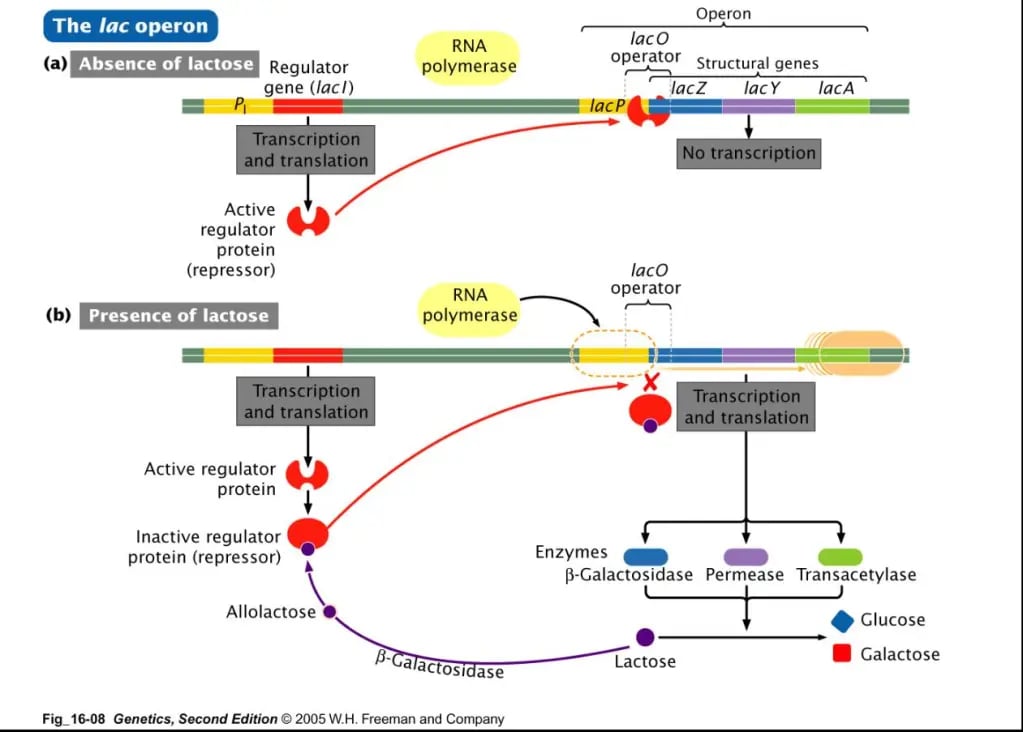 When lactose is present
When lactose is present
When lactose is present in a medium, some of it will be converted to allolactose. This allolactose binds to the repressor protein. Binding of allolactose with repressor protein changes the shape of repressor protein so it can no longer binds to the operator region. This allows the RNA polymerase to bind to the promoter site, starting the initiation of transcription of the structural genes lacZ, lacY, and lacA to produce mRNA.
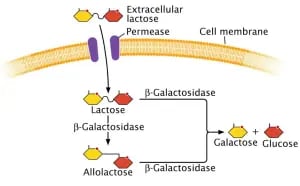 Once the mRNA is produced, it is translated to produce enzymes. The lacZ gene encodes for the production of the enzyme β-galactosidase (B), lacY gene for the production of the enzyme permease (P) and the lacA gene is responsible for the production of the enzyme thiogalactoside transacetylase.
Once the mRNA is produced, it is translated to produce enzymes. The lacZ gene encodes for the production of the enzyme β-galactosidase (B), lacY gene for the production of the enzyme permease (P) and the lacA gene is responsible for the production of the enzyme thiogalactoside transacetylase.
Catabolite repression (glucose effect)
Enzyme for lactose utilization can be induced by adding lactose in the growth medium but, it would be wasteful to induce these enzymes if the cells are already growing on a carbon source that they could use more efficiently e.g. glucose. This regulation is governed by a phenomenon called catabolite repression, also known as glucose effect. Catabolite repression ensures that the cells use the BEST carbon source first.
In the case of catabolite repressible enzymes, binding of RNA polymerase in the promoter region of DNA occurs only if catabolite activator protein (CAP) also known as CRP (cAMP Receptor Protein) has bound first. The gene encoding CAP is located elsewhere on the bacterial chromosome, not linked to the lac genes. CAP is an allosteric protein which binds to DNA only if it has first bound with cyclic AMP.
Cyclic AMP is synthesized from ATP by an enzyme called adenylate cyclase, and glucose inhibits the synthesis of cyclic AMP and stimulates its transport out of the cell.
When glucose is transported into the cell, the cyclic AMP level in the cell is lowered. This means only few CAP (which were bound with cyclic AMP) will be able to bind to DNA. This will cause loose binding of RNA polymerase to the promoter region resulting low level of transcription. When there is no any cyclic AMP left in the cell, the transcription will be ceased.
High glucose in the medium
When glucose levels are high, no cAMP is made, the CAP–cAMP complex does not form. CAP cannot bind DNA without cAMP and RNA polymerase cannot bind to the promoter efficiently. In this case, transcription occurs only at a low level.
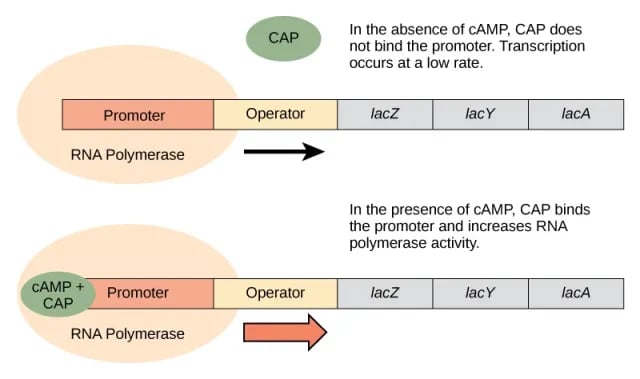 Figure: Expression of the lac genes in the presence of Glucose (Image source-Ref.1)
Figure: Expression of the lac genes in the presence of Glucose (Image source-Ref.1)
Low glucose in the medium
When the level of glucose in the environment is low or nil, abundant cAMP binds CAP to form the CAP-cAMP complex, which binds DNA. When CAP–cAMP binds DNA, the efficiency of RNA polymerase binding is increased at the lac operon promoter resulting in a higher level of transcription of the structural genes.
Regulation of Lac Operon
For the transcription of structural genes by the lac operon, two requirements must be met:
- Glucose must be absent: The level of cyclic AMP must be high enough so that the CAP protein binds to the CAP binding site. Bound CAP helps to attach RNA polymerase efficiently to the lac operon promoter.
- Lactose must be present: There must be an inducer (such as lactose) so that lactose repressor does not block transcription by binding to the operator.
Each of the regulatory proteins (CAP and lac repressor) responds to one environmental signal and communicates it to the lac genes. The combined effect of these two regulators ensures that the genes are expressed at significant levels only when lactose is present and glucose is absent. Now, let’s observe the transcription of the operon in various environmental conditions:
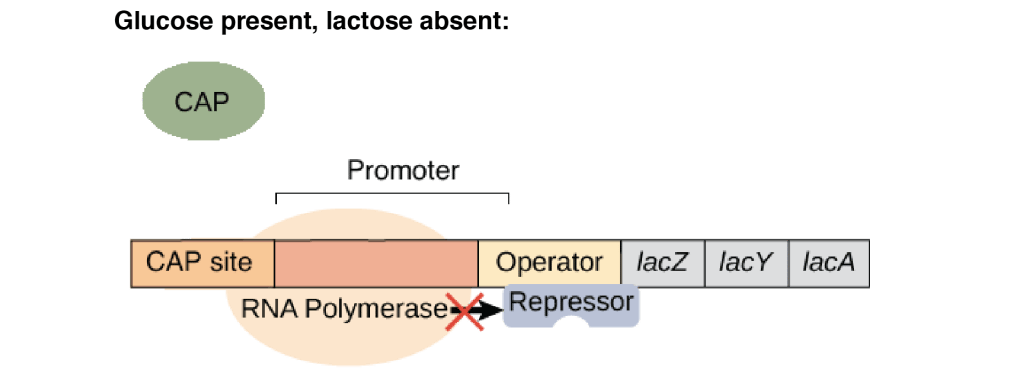
1. Glucose present, lactose absent
As glucose is present, cAMP level is low so activator CAP remains inactive. Lac repressor remains bound to the operator and prevents binding of RNA polymerase. In this condition, no transcription of the lac operon occurs.
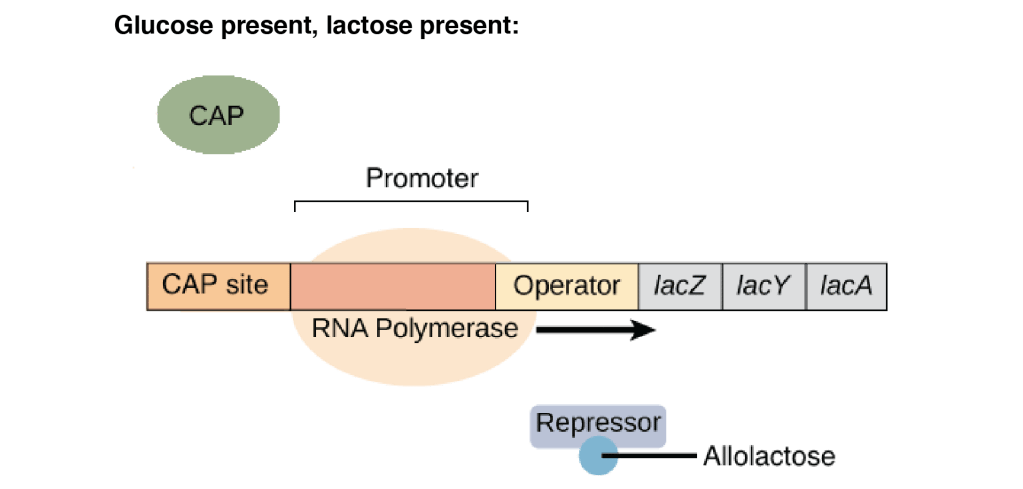
 2.Glucose present, lactose present
2.Glucose present, lactose present
Activator CAP remains inactive. The lac repressor is not functional because the inducer (lactose) is present. In this condition, the basal level transcription of the lac operon occurs.
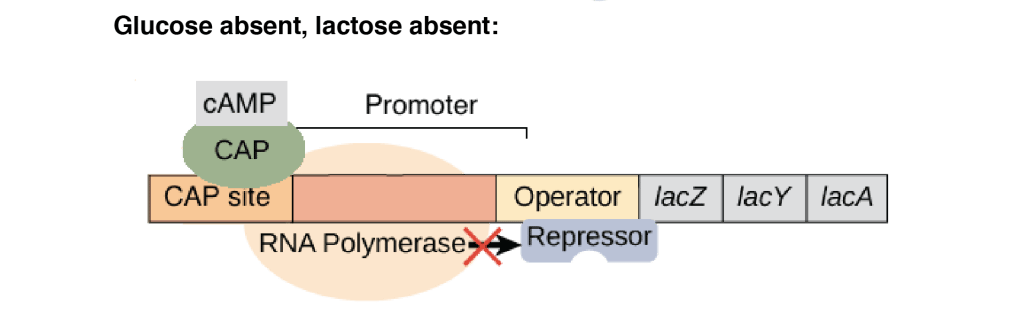
 3.Glucose absent, lactose absent
3.Glucose absent, lactose absent
Activator CAP is active as a high level of cAMP is present (as glucose is absent) but lac repressor is functional (active). Lac repressor remains bound to the operator and prevents transcription.
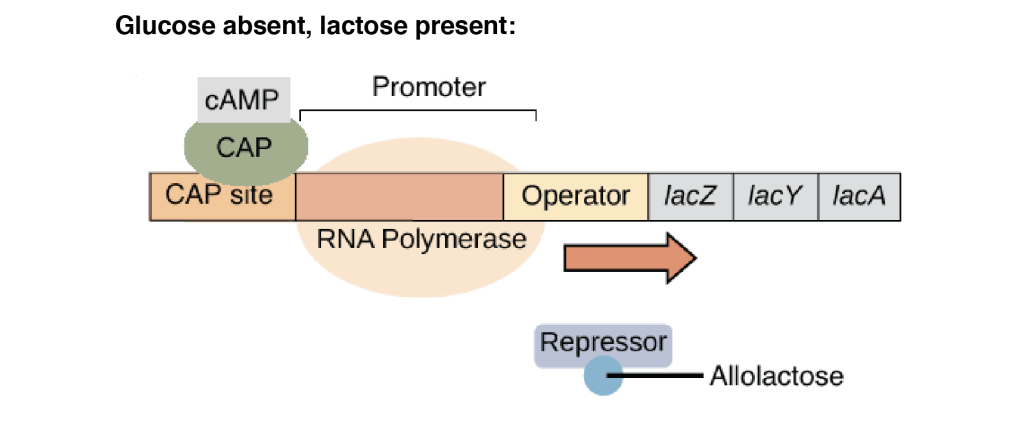
 4.Glucose absent, lactose present
4.Glucose absent, lactose present
cAMP levels are high so CAP is active and bound to the DNA. CAP helps the efficient binding of RNA polymerase to the promoter. lac repressor is inactive due to the presence of inducer (lactose/allolactose). In this condition, strong transcription of the lac operon occurs.
 Summary of Lac Operon Responses
Summary of Lac Operon Responses
| Glucose | Binding of CAP | Lactose | Binding of Repressor | Level of Transcription |
|---|---|---|---|---|
| + | – | – | + | No transcription |
| + | – | + | – | Low level transcription |
| – | + | – | + | No transcription |
| – | + | + | – | High level transcription |
References and further reading
- Prokaryotic gene regulation by OpenStax College, Biology (Images showing effects of Glucose and Lactose in lac operon were modified from “Prokaryotic gene regulation: Figure 3“
- Molecular Biology of the Gene (5th Edition), by James D. Watson
- Brock Biology of Microorganisms(15th Edition) by Michael T. Madigan
{kind=link}

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.