Macrolides: Mode of Action, Mechanism of Resistance
Macrolides: Mode of Action, Mechanism of Resistance
Macrolides are primarily bacteriostatic antibacterial agents that inhibit bacterial protein synthesis by binding with 50S subunit of the ribosome. Members of the group include erythromycin, azithromycin, clarithromycin, roxithromycin, dirithromycin, and telithromycin.
These were originally derived from Saccharopolyspora erythraea (originally called Streptomyces erythreus), a type of soil-borne bacteria.
Erythromycin was the first macrolide discovered and first used in 1952 as a substitute to penicillin in cases where patients were allergic to penicillin or had penicillin-resistant illnesses.
These are effective against gram-positive (excluding enterococci) but are not effective against most genera of gram-negative bacteria because of uptake difficulties associated with gram-negative outer membranes. They are also active against Mycoplasma pneumoniae, Treponema pallidum, Bordetella pertussis, Chlamydia trachomatis, Chlamydophila pneumoniae, Legionella spp., Campylobacter spp., and Borrelia spp.
Structure
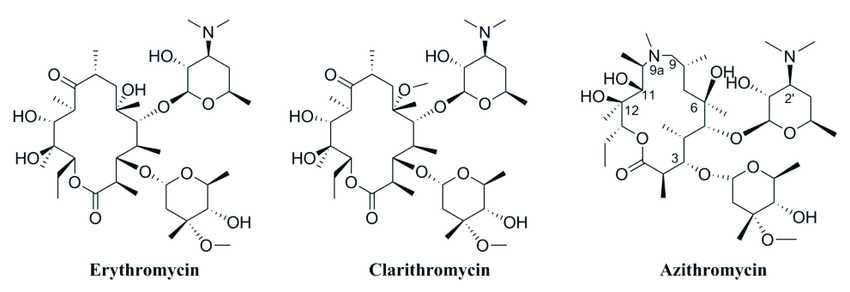 Figure: Structure of macrolides
Figure: Structure of macrolides
Structurally, these consist of a large macrocyclic lactone ring to which one or more deoxy sugars or amino sugar residues are attached.
Macrolide antibiotics are classified according to the size of the macrocyclic lactone ring (either 12‐, 14‐, 15‐ or 16‐membered ring) and sources (natural or synthetic).
Mode of Action of Macrolides
The macrolides bind to the 50S ribosomal subunit with a specific target in 23s ribosomal RNA molecule and various ribosomal proteins. All macrolides inhibit bacterial protein synthesis, however, different macrolide act at different steps.
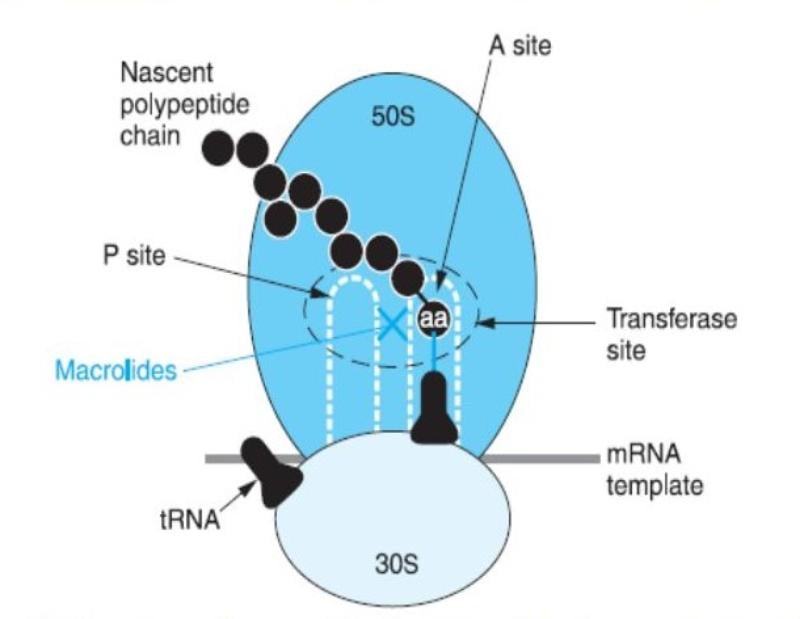 Figure: Mode of action of Macrolides
Figure: Mode of action of Macrolides
The 14-membered macrolides block the translocation of peptidyl-tRNA and the 16-membered compounds inhibit the peptidyl transfer reaction. The most recent hypothesis suggests that all macrolides stimulate dissociation of peptidyl-tRNA from the ribosomes during the elongation phase, leading to the inhibition of protein synthesis.
Mechanism of Resistance
Resistance to macrolides occurs by two different mechanisms:
Altered target
Enzymatic alteration of ribosomal target reduces drug binding.
Mutations in 23S rRNA, L4, and/or L22 ribosomal proteins can confer macrolide resistance because the mutation is technically in the 23S rRNA gene. Erm genes that encode rRNA methyltransferases mediates a major and widespread mechanism of resistance to the macrolide class of antibiotics by adding one or two methyl groups to the exocyclic amino group located in 23S rRNA.
Enzymatic detoxification of the drug
These can be inactivated by enzymes such as esterases or phosphotransferases, or by glycosyltransferases, or formyl reductases encoded in plasmids of both gram-negative and gram-positive bacteria.
Active efflux of the drug
There are two families of macrolide efflux pumps, transcriptionally mediated—mef, a major-facilitator-superfamily pump that confers resistance to most 14- and 15-membered macrolides and msr, a member of the ATP-binding cassette (ABC) superfamily that generally confers resistance to 14- and 15-membered macrolides and streptogramin B and low-level resistance to ketolides.
References and further readings
- Bryskier, A., & Bergogne-Bérézin, E. (2005). Macrolides. Antimicrobial Agents, 475–526.
- Fyfe, C., Grossman, T. H., Kerstein, K., & Sutcliffe, J. (2016). Resistance to Macrolide Antibiotics in Public Health Pathogens. Cold Spring Harbor Perspectives in Medicine, 6(10).
- Mazzei, T., Mini, E., Novelli, A., & Periti, P. (1993). Chemistry and mode of action of macrolides. Journal of Antimicrobial Chemotherapy, 31(suppl_C), 1–9.

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.