Bacterial Conjugation and F Plasmid Transfer: Mechanism, Terminology, and Role in Antibiotic Resistance
How a bacterial "mating bridge" moves a fertility plasmid from donor to recipient, the 1946 experiment that proved bacteria have sex at all, and why this exact mechanism spreads ESBL and carbapenemase resistance today.
Proving that bacteria have sex, then proving they need to touch to do it
In 1946, Joshua Lederberg and Edward Tatum ran a deceptively simple experiment. They had two strains of E. coli K-12, each with a different set of nutritional deficiencies. Strain A could make its own threonine, leucine, and thiamine, but couldn't make methionine or biotin, so it needed those supplied externally to grow. Strain B was the exact opposite: it could make methionine and biotin, but not threonine, leucine, or thiamine.
Neither strain, grown alone on a minimal medium with no supplements, could grow at all. But when Lederberg and Tatum mixed the two strains together first, then plated the mixture on that same bare minimal medium, colonies appeared. Some bacteria in the mix had somehow gained the ability to make everything they needed, a genetic capability neither parent strain possessed on its own.
Genetic material had clearly moved between the two strains. But how? A few years later, Bernard Davis designed a way to find out: a U-shaped glass tube with a filter down the middle, fine enough to block whole bacterial cells from crossing but not much else. When the two E. coli strains were grown on opposite sides of that filter, with no direct contact possible, no recombinant colonies appeared at all. Whatever had happened in the original experiment, it required actual physical contact between living cells.
That requirement, direct cell-to-cell contact, is what defines conjugation, and it's also the single clearest way to tell it apart from the other two gene transfer mechanisms in this cluster: transformation needs no contact at all, and transduction (discovered a few years later using this exact same U-tube trick) turned out not to need contact either, since a phage can carry DNA straight through a filter that would stop a conjugating bacterium cold.
The vehicle for that contact-dependent transfer is usually a plasmid, and the best-studied example, the one Lederberg and Tatum's experiment ultimately traced to, is the F plasmid. The same basic transfer mechanism it uses is, today, one of the primary ways ESBL- and carbapenemase-producing genes spread silently between E. coli and Klebsiella pneumoniae in hospitals.
Conjugation in bacteria is one of the three processes of horizontal gene transfer, alongside transformation and transduction. It was the first of the three to be extensively studied. Conjugation occurs strictly between two living cells, requires direct cell-to-cell contact, and involves the mobilization of either a plasmid or, in some strains, a portion of the donor cell's chromosome.
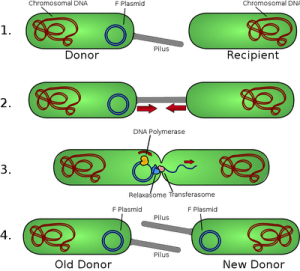 Figure: Bacterial conjugation and transfer of F plasmid
Figure: Bacterial conjugation and transfer of F plasmid
Conjugative plasmids transfer themselves between bacteria, and this exact mechanism has driven much of the spread of antibiotic resistance among pathogenic bacteria.
Key plasmid background
Before the mechanism itself, a few facts about plasmids in general:
- Plasmids are small, circular pieces of DNA, separate from and replicating independently of the bacterial chromosome.
- A plasmid can carry genes for specialized functions, most importantly for this topic, drug resistance.
- Plasmids may encode the genes needed to mediate their own transfer to another cell, but not every plasmid is capable of conjugative transfer.
- Plasmids capable of transferring themselves independently are called self-transmissible (conjugative) plasmids; those that cannot are non-transmissible.
F plasmid terminology
The F plasmid, also called the fertility factor, confers donor characteristics (the sex pilus) on the bacterial cell carrying it.
| Strain | Definition |
|---|---|
| F+ strains | Bacteria carrying the F plasmid; referred to as male/donor |
| F– strains | Bacteria lacking the F plasmid; referred to as female/recipient |
| Hfr strains | Arise from F+ strains when the F plasmid integrates into the bacterial chromosome at one of several possible sites (see the Conjugation: Transfer of Chromosomal DNA by HFr Strains article for the full Hfr mechanism) |
| F′ (F-prime) strains | Arise when an integrated F plasmid excises imprecisely from an Hfr chromosome and carries a fragment of chromosomal DNA out with it |
Mechanism of F plasmid transfer by conjugation
- Contact. The F plasmid alters the donor cell's surface, producing a sex pilus that establishes contact with an F– recipient cell.
- Mating bridge formation. The sex pilus draws the two cells together and retracts, forming a stable conjugation (mating) bridge, a temporary cytoplasmic connection between donor and recipient.
- Nicking at oriT. An enzyme called relaxase nicks one strand of the F plasmid at a specific site called the origin of transfer (oriT).
- Single-strand transfer. The nicked strand is fed through the conjugation bridge into the recipient cell, 5′ end first, while the donor cell simultaneously synthesizes a new complementary strand to replace the one leaving, a process called rolling circle replication.
- Complementary strand synthesis in the recipient. Once inside the recipient, the transferred single strand is used as a template to synthesize its own complementary strand, recircularizing into a complete, double-stranded F plasmid.
- New donor created. The recipient cell now carries a complete copy of the F plasmid and itself becomes an F+ donor cell, capable of transferring the plasmid onward to further F– recipients.
Because the donor retains its own copy of the plasmid throughout this process (rolling circle replication continuously regenerates it), mixing F– and F+ cells in the same culture rapidly converts the entire population to F+ over just a few generations.
Why This Matters Clinically
- ESBL and carbapenemase spread. Conjugative plasmids are the primary vehicle by which extended-spectrum beta-lactamase (ESBL) and carbapenemase genes spread between Escherichia coli, Klebsiella pneumoniae, and other Enterobacterales, both within a single patient's gut flora and between patients in a hospital setting. This is the same F-plasmid-style mechanism described above, just carrying resistance genes instead of (or in addition to) fertility genes.
- Shigella-mediated dysentery. Transfer of R (resistance) plasmids between Shigella species by conjugation has been directly linked to the emergence of antibiotic-resistant, dysentery-causing strains.
- Cross-kingdom transfer. Conjugation isn't limited to bacterium-to-bacterium transfer. Agrobacterium tumefaciens uses a conjugation-like mechanism to transfer the T-DNA portion of its Ti (tumor-inducing) plasmid directly into plant cells, causing crown gall tumors, a striking demonstration of how broadly this transfer machinery can operate, even though it is a plant pathogen rather than a human one.
How to Remember
The bacterial family-tree mnemonic for F+/F–/Hfr/F′:
- F+ has the plasmid, free-floating, and is the donor
- F– lacks the plasmid entirely and is the recipient
- Hfr is an F+ cell whose plasmid has settled down by integrating into the chromosome
- F′ is an Hfr cell whose plasmid packed its bags carelessly on the way back out, taking a piece of chromosome with it
The needle-and-thread picture for rolling circle transfer: picture the F plasmid as a circular thread. The donor nicks the thread at one fixed point (oriT) and feeds the loose end through the mating bridge like thread through a needle, into the recipient, while simultaneously spinning up a brand-new copy of the thread to replace the one leaving. Neither cell ever ends up without a copy.
Anchor for why F+ × F– rapidly converts a whole population: since the donor never loses its plasmid and the recipient gains a full copy every single time, mixing F+ and F– cells is like a chain letter that never runs out of copies to send. Within a few generations, essentially everyone in the tube has become F+.
Anchor for the clinical stakes: the same rolling-circle handoff that turns an F– cell into an F+ cell in a test tube is, in a hospital gut flora sample, the mechanism handing a carbapenemase gene from one Klebsiella to the next.
Key exam facts in one table
| Fact | Detail |
|---|---|
| Definition | Direct, contact-dependent transfer of a plasmid (or, in Hfr strains, chromosomal DNA) from a donor to a recipient bacterium |
| Requires cell contact? | Yes, always, via a sex pilus and conjugation bridge — the defining difference from transformation and transduction |
| Discovered by | Lederberg and Tatum (1946), using auxotrophic E. coli K-12 strains; contact requirement confirmed by Davis's U-tube experiment |
| Fertility factor | The F plasmid, which confers the sex pilus and donor phenotype |
| F+ | Has the F plasmid (donor) |
| F– | Lacks the F plasmid (recipient) |
| Hfr | F plasmid integrated into the chromosome; "high frequency of recombination" |
| F′ | Hfr-derived plasmid carrying a fragment of chromosomal DNA |
| Site of nicking | Origin of transfer (oriT), cut by relaxase |
| Replication mechanism | Rolling circle replication in the donor while a single strand transfers to the recipient |
| Outcome of F+ × F– mating | Recipient becomes F+; donor retains its own copy; whole population converts to F+ over time |
| Clinical relevance | Major mechanism for spread of ESBL and carbapenemase genes among Enterobacterales; R-plasmid spread in Shigella |
Where Students Get Confused
- "Conservative" gene transfer vs. conservative DNA replication. The original description of conjugation as a "conservative type of gene transfer" (because the donor keeps a copy) is easy to confuse with the conservative model of DNA replication from the Meselson–Stahl experiment, which is a completely different concept (and one that was disproven in favor of semiconservative replication). Here, "conservative" just means the donor doesn't lose its plasmid, nothing to do with how DNA strands are copied.
- F+ × F– mating vs. Hfr × F– mating. This is the most consequential mix-up in the whole conjugation topic. F+ × F– conjugation transfers only the plasmid, is fast, and converts the recipient to F+. Hfr × F– conjugation transfers chromosomal genes, is much slower, and the recipient typically does not become F+ (see the Hfr article for the full explanation).
- Conjugative vs. mobilizable vs. non-transmissible plasmids. A conjugative plasmid can transfer itself independently. Some plasmids are "mobilizable," meaning they can be transferred only with help from a separate conjugative plasmid's machinery. Non-transmissible plasmids cannot transfer by conjugation at all.
- Assuming conjugation always transfers the whole plasmid intact as double-stranded DNA. Only a single strand crosses the mating bridge; each cell then synthesizes its own complementary strand independently.
References
- Snustad, D., & Simmons, M. (2012). Principles of Genetics (6th ed., pp. 175–179). Hoboken, NJ: Wiley.
- Watson, J., Baker, T., Bell, S., Gann, A., Levine, M., & Losick, R. (2004). Molecular Biology of the Gene (5th ed., pp. 655–658). San Francisco: Pearson Education, Inc.
- Lederberg, J., & Tatum, E. L. (1946). Gene recombination in Escherichia coli. Nature, 158(4016), 558.
- Davis, B. D. (1950). Nonfiltrability of the agent of genetic recombination in Escherichia coli. Journal of Bacteriology, 60(4), 507–508.
Frequently Asked Questions
What is bacterial conjugation?
What is the F plasmid?
What is the difference between F+, F-, Hfr, and F' strains?
Who discovered bacterial conjugation?
How does the F plasmid actually move between cells?
Why does mixing F+ and F- bacteria convert the whole population to F+?
How is conjugation different from transformation and transduction?
Why is bacterial conjugation clinically important?

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.