Influenza (flu) Virus: Classification, Structure, and Diagnosis
Why influenza mutates faster than immunity can keep up, how drift differs from shift, and why your flu vaccine needs updating every year.
In 1918, an influenza pandemic killed somewhere between 50 and 100 million people worldwide, more than the First World War. What made the 1918 strain so destructive was not unusual virulence alone but the fact that it carried surface proteins that no living person's immune system had ever encountered before. There was no pre-existing immunity, no vaccine, and no antiviral treatment. The virus swept through populations that were completely defenceless against it.
A century later, influenza still kills between 290,000 and 650,000 people annually worldwide, even in seasons when pandemic strains aren't circulating. The reason is the same biological principle that made 1918 possible: the influenza virus mutates faster than the human immune system can keep up with it. Immunity built against this year's strain offers only partial protection against next year's, because the virus you encounter next flu season will not look quite the same to your immune cells as the one you saw this year.
Two molecular mechanisms drive this. The first is slow and continuous; gradual mutation accumulating in the surface proteins year over year. The second is sudden and catastrophic; wholesale exchange of genetic segments between two different influenza strains co-infecting the same cell, capable of producing a completely new virus with surface proteins that no living human has ever been exposed to. Understanding both mechanisms is what makes influenza clinically coherent: it explains why you need a new flu vaccine every year, why certain pandemic strains appear with little warning, and why neuraminidase inhibitors work the way they do.
Influenza, commonly called “the flu,” is a contagious respiratory illness, a very important global public health problem. It causes seasonal flu epidemics yearly and is a critical disease responsible for school and job absenteeism, hospitalization, and deaths. Flu pandemics have killed millions of people worldwide; another flu pandemic is looming in our future.
Influenza (flu) is caused by influenza viruses, a member of the orthomyxovirus family. Influenza viruses infect the respiratory tract (i.e., nose, throat, lungs) and can cause mild to severe life-threatening illnesses. There are three types of influenza (flu) viruses: A, B, and C. Antigenic differences exhibited by two internal structure proteins (NP and M) are used to divide influenza viruses into types A, B, and C.
Influenza viruses A and B cause seasonal epidemics, whereas type C causes mild respiratory illness. Influenza B viruses have historically circulated as two lineages, B/Yamagata and B/Victoria. B/Yamagata is considered effectively eliminated from global circulation following its absence since 2020; the quadrivalent influenza vaccines used from 2024 onwards have moved to trivalent formulations excluding B/Yamagata.
Influenza A virus has been further divided into subtypes and strains. The mutability and high frequency of genetic reassortment in this virus result in antigenic changes in viral surface glycoprotein, making influenza type A antigenically highly variable and responsible for most cases of epidemic influenza.
Subtypes of Influenza A viruses
Influenza A viruses are divided into subtypes based on two proteins on the virus’s surface: hemagglutinin (H) and neuraminidase (N). There are 18 different hemagglutinin subtypes (H1 through H18) and 11 neuraminidase (N1 through N11) subtypes.
Strains of Influenza A viruses
Influenza A viruses can be further broken down into different strains. Some of the most successful and virulent strains of flu are
- H5N1 avian (bird) flu: caused outbreaks in domestic poultry in parts of Asia and the Middle East. Human infection with H5N1 is rare, though case numbers have risen since 2020 with ongoing outbreaks in poultry and sporadic human exposure events.
- H1N1 (swine flu): caused a worldwide pandemic in 2009.
- Influenza A H3N2 variant viruses
Nomenclature (naming) of Influenza Virus
Internationally accepted naming convention is followed for the naming of influenza viruses. The approach uses the following components:
- The antigenic type (e.g., A, B, C)
- The host of origin (e.g., swine, chicken, duck, etc. For human-origin viruses, no-host of origin designation is given.)
- Geographical origin (e.g., Hong Kong, Taiwan, etc.)
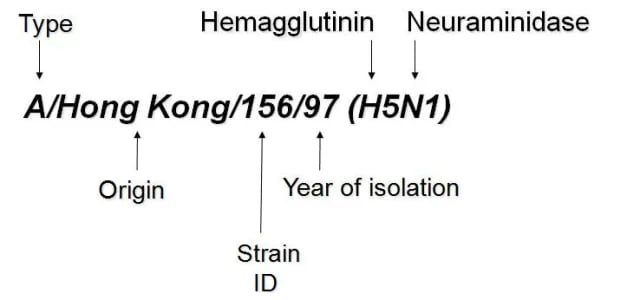 Figure: Fig: Naming of influenza virus
Figure: Fig: Naming of influenza virus
Strain number (e.g., 15, 7, etc.) Year of isolation (e.g., 57, 2009, etc.) For influenza, A viruses, the hemagglutinin, and neuraminidase antigen descriptions are in parentheses (e.g., (H1N1), (H5N1)
For example:
- A/duck/Alberta/35/76 (H1N1) for a virus of duck origin
- A/Perth/16/2009 (H3N2) for a virus of human origin
Structure of Influenza virus
- Influenza virions are usually spherical(diameter 80-110 nm).
- An enveloped virus, the outer layer is a lipid membrane that is taken from the host cell.
- Inserted into the lipid membrane are ‘spikes,’ about 10 nm long, glycoproteins are known as HA (hemagglutinin) and NA (neuraminidase).
# NA is a target of antiviral drugs called neuraminidase inhibitors, including oseltamivir (Tamiflu) and zanamivir (Relenza), which block viral release from infected cells- see the Treatment section below. - HA and NA are the important antigens that determine
antigenic variation of influenza viruses and host immunity. subtype and strains of influenza virus (A/H1N1, for example).
- Protection against re-infection is mainly due to developing antibodies to HA (but antibodies to NA are also protective).
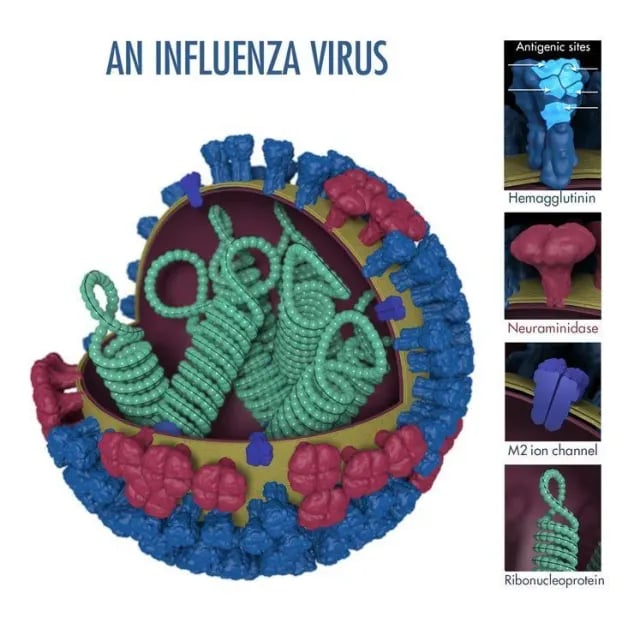 Figure: Influenza (flu) virus structure
Figure: Influenza (flu) virus structure
Image source: CDC
- M2 proteins are also embedded in the lipid membrane. M2 proteins are the targets of antiviral drugs adamantanes – amantadine and rimantadine.
- Beneath the lipid membrane is a viral protein called M1, or matrix protein. Matrix protein is a major component of the virion (about 40% of viral protein), forms a shell, and gives strength and rigidity to the lipid envelope.
- Within the interior of the virion are the genetic material of the virus; the viral single-stranded, negative-sense, viral RNAs
Eight of them for influenza A and B viruses; the influenza C virus has only seven segments; lacking a neuraminidase gene); Most of the segments code for a single protein.
- Each RNA segment consists of RNA joined with several proteins PB1, PB2, PA, and NP (RNA Polymerase), responsible for RNA transcription and replication.
- The interior of the virion also contains another protein called NEP.
Pathogenesis of Influenza Infection
Entry and initial replication
Influenza virus is transmitted via respiratory droplets and aerosols generated when an infected person talks, coughs, or sneezes. Inhalation deposits the virus on the epithelial surface of the upper respiratory tract — primarily the nasal mucosa and pharynx — where it binds to sialic acid residues on the surface of respiratory epithelial cells via the hemagglutinin (HA) glycoprotein.
The specificity of HA binding to different forms of sialic acid linkage is a key determinant of host range:
- Human-adapted influenza viruses preferentially bind α-2,6-linked sialic acid, which is expressed on epithelial cells of the upper human respiratory tract
- Avian influenza viruses preferentially bind α-2,3-linked sialic acid, found in the avian gut and also in the human lower respiratory tract
This receptor preference explains why most human-adapted flu viruses cause upper respiratory disease (binding occurs easily at the nose and pharynx), while avian influenza strains that do infect humans (like H5N1) tend to cause more severe lower respiratory tract disease — they can only efficiently bind the deep lung cells that express α-2,3 linkages.
Viral replication and spread
After HA-mediated fusion and entry, the eight RNA segments are transcribed and replicated in the nucleus using the viral RNA-dependent RNA polymerase (PB1, PB2, PA complex). New virions assemble and bud from the cell surface. Neuraminidase (NA) cleaves sialic acid bonds that would otherwise trap budding virions on the cell surface, releasing them to infect adjacent cells. This is the precise step targeted by neuraminidase inhibitors (see Treatment section).
Influenza replication is rapid: new virions can be produced within 6 hours of infection, and peak viral shedding occurs within the first 24–72 hours of symptoms, often before the infected person knows they are sick.
Immune response and why some patients develop severe disease
The immune response to influenza is predominantly mediated by CD8+ cytotoxic T cells (which kill infected epithelial cells) and neutralising antibodies against HA and NA. The local mucosal and systemic inflammatory response produces the characteristic systemic symptoms of influenza — fever, myalgia, and malaise — which are largely cytokine-driven rather than directly caused by the virus itself.
Severe disease (pneumonia, respiratory failure) occurs when viral replication is not contained in the upper respiratory tract and spreads to the lower respiratory tract and alveoli. Risk factors for severe disease reflect the host's ability to mount an effective early immune response:
- Elderly patients: declining immune function and reduced mucosal defences
- Young children: immature immune systems, first exposure to influenza
- Pregnant women: altered immune regulation during pregnancy
- Immunocompromised patients: reduced T-cell and antibody responses
- Obese individuals: adipose tissue dysfunction and impaired immune signalling
- Patients with chronic cardiopulmonary disease: less respiratory reserve if pneumonia develops
Clinical Features of Influenza
Typical uncomplicated influenza
The incubation period is 1–4 days (average 2 days). The onset of influenza is characteristically abrupt, distinguishing it from the common cold, which develops gradually:
- Sudden high fever (38–40°C), chills
- Severe headache
- Pronounced myalgia and malaise (often described as "hit by a truck")
- Dry cough, sore throat, nasal congestion
- Loss of appetite
Systemic symptoms typically dominate in the first 2–3 days; respiratory symptoms persist longer. In uncomplicated influenza, symptoms resolve within 7 days in most healthy adults, though fatigue may persist for 1–2 weeks.
Complications
Primary viral pneumonia: Influenza virus itself extends into the lower respiratory tract, causing diffuse bilateral pneumonia. More common with pandemic strains, H5N1, and in immunocompromised patients.
Secondary bacterial pneumonia: The most common serious complication. Influenza disrupts mucociliary clearance and impairs local immune defences, creating ideal conditions for Streptococcus pneumoniae, Staphylococcus aureus (including MRSA), and Haemophilus influenzae to establish lower respiratory infection. Many of the deaths in the 1918 pandemic were caused by secondary bacterial pneumonia, not the virus itself.
Reye's syndrome: Acute hepatic failure and encephalopathy in children given aspirin during influenza (or varicella) infection. Aspirin must not be given to children with influenza. Paracetamol or ibuprofen should be used instead.
Myocarditis and pericarditis: Rare but recognised complications, particularly in young adults with severe influenza.
Antigenic Variation: Why Influenza Evades Immunity
Influenza's ability to return year after year, and occasionally as pandemic strains no living person has immunity to, depends on two distinct genetic mechanisms. Understanding both — and the difference between them — is one of the highest-yield topics in influenza biology.
Antigenic drift: the slow, annual change
The influenza RNA polymerase (the PB1/PB2/PA complex) is error-prone: it lacks the proofreading mechanism that cellular DNA polymerases use to correct copying mistakes. Every replication cycle produces progeny viruses with slight genetic differences from the parent — point mutations accumulating randomly across the genome.
Most of these mutations are neutral or harmful to the virus and those virions don't survive. But mutations that fall within the antigenic sites of HA or NA — the regions that stick out on the surface and are directly targeted by neutralising antibodies — can change the shape of those proteins just enough that antibodies from a previous infection or vaccination no longer recognise them effectively. When such a mutation confers a selective advantage (the virus can now infect previously immune hosts), it spreads through the population.
This gradual, mutation-by-mutation accumulation of surface protein changes is called antigenic drift. It happens continuously, in every flu season, in every geographic location where influenza is circulating. The practical consequence: the influenza strain circulating in 2027 will look slightly different to the immune system than the strain from 2026, which is why flu vaccines must be reformulated annually to match the currently circulating strains.
Drift is unpredictable in its precise direction — the mutations are random — which is why WHO laboratories monitor circulating strains year-round to make the best possible prediction of which variants to include in the coming season's vaccine.
Antigenic shift: the sudden, pandemic-generating change
While drift changes surface proteins gradually, antigenic shift replaces them wholesale. Shift is only possible because of influenza's segmented genome: its eight RNA segments are packaged separately inside the virion, not as one continuous strand.
If two different influenza A strains infect the same host cell simultaneously — for example, a human-adapted H3N2 strain and an avian H5N1 strain — both sets of eight RNA segments are present in the same cellular pool. During virion assembly, the eight segments are packaged somewhat randomly. A new hybrid virus can emerge carrying, for example, the HA segment from the avian strain and most other segments from the human strain. This reassortment produces a virus with surface proteins that are completely new to the human immune system — not just slightly changed, but entirely unfamiliar.
Such a reassortant can spread explosively through a globally susceptible population because no one has prior immunity to its surface proteins. This is the mechanism behind influenza pandemics:
| Year | Pandemic strain | Mechanism |
|---|---|---|
| 1918 | H1N1 ("Spanish flu") | Novel H1 antigen, estimated 50–100 million deaths |
| 1957 | H2N2 ("Asian flu") | Reassortment introducing new H2/N2 antigens |
| 1968 | H3N2 ("Hong Kong flu") | Reassortment introducing new H3 antigen |
| 2009 | H1N1 ("Swine flu") | Reassortment of human, avian, and swine gene segments |
Why does reassortment most often involve pigs? Pigs express both α-2,6-linked sialic acid (the human-adapted receptor) and α-2,3-linked sialic acid (the avian receptor) on their respiratory epithelial cells. This dual receptor expression allows pigs to be co-infected by both human-adapted and avian influenza strains simultaneously — creating the cellular environment where reassortment between the two lineages can occur. Pigs have therefore historically been considered "mixing vessels" where pandemic strains can emerge.
Key difference between drift and shift:
| Feature | Antigenic drift | Antigenic shift |
|---|---|---|
| Mechanism | Point mutations in HA/NA genes | Reassortment of entire RNA segments |
| Speed | Gradual, continuous | Sudden, unpredictable |
| Scale | Annual epidemics | Pandemics |
| Influenza types | A, B | A only (B is mainly human-restricted, limited reassortment) |
| Immunity gap | Partial (some cross-reactivity from prior immunity) | Complete (no prior immunity in population) |
Laboratory Diagnosis
For the laboratory diagnosis of influenza virus infection, specimens should be taken as early as possible during the disease, preferably within the first 72 hours after the onset of symptoms. Routine specimens for influenza virus diagnosis include
- Nasal washings
- Nasopharyngeal aspirate
- Nasopharyngeal swab
- Nasal and throat swabs
For successful isolation, a specimen should be placed on a viral transport medium to stabilize the virus
Cell Culture
Madin-Darby kidney cell Line (MDCK) supplemented with trypsin supports growth and multiple cycles of influenza A, B, and C. Standard virus isolation is done in cell culture tubes seeded with MDCK cells. The cultures are incubated at 34°C in a stationary rack or roller machine. Some influenza viruses cause a distinct cytopathic effect in MDCK cells several days after inoculation. Negative cultures should be checked by a hemadsorption or hemagglutination test at 2-3 days intervals.
Detection of Infected Cells in Clinical Specimens by Immuno-fluorescence
Demonstration of infected epithelial cells in nasopharyngeal aspirate or nasopharyngeal swab is a sensitive method (50-90% more sensitive than virus isolation) for the diagnosis of influenza. The result can be obtained within a few hours after the specimen collection. Exfoliated cells in NPA and NPS are prepared, applied to microscopic slides, and fixed. Polyclonal sera and/or monoclonal antibodies are used as a reagent for the staining process.
Detection of Influenza Virus Antigens by Immunoassays
Various tests such as radioimmunoassay, enzyme immunoassay (EIA), and fluoroimmunoassay have been developed to detect influenza virus antigens directly in clinical specimens or after amplification in cell culture. Antigens in clinical samples or cell culture material are commonly captured by specific antibodies that have previously adsorbed to the solid phase. After incubation, unbound material is washed, the bound antigen is reacted with a secondary antibody and then incubated further with a labeled anti-species antibody.
Reverse Transcriptase PCR
The reverse transcriptase PCR has been applied to detect influenza virus in clinical specimens and cell culture or egg-grown material. The viral RNA is first transcribed into DNA, then amplified in a second step using suitable primers and DNA Polymerase. Amplified DNA is electrophoresed in an agarose gel, and DNA is detected by staining with ethidium bromide. Molecular weight markers are included to identify the amplified DNA of the appropriate size. DNA can also be detected by molecular hybridization with specific labeled probes either in Southern blots or by spot hybridization. Appropriate selection of primers permits type-specific identification of influenza virus A, B, and C or subtype-specific identification.
Serology
Although serologic methods seldom yield a result early enough to influence the patient’s treatment, they often establish the diagnosis of influenza virus infection when other methods do not detect the virus. When the sensitivities of newly developed diagnostic tests are being evaluated, serology should be considered instead of or in combination with virus isolation as the “Gold Standard.” Hemagglutination inhibition test (HAI), the neutralization test (NT), and the complement fixation test (CFT) are traditional methods in serodiagnosis and sero-epidemiologic studies of influenza, but during recent years, EIA has found wide application. The complement fixation test measures antibodies against the NP and thus allows the type-specific detection of antibodies to influenza A or B viruses. HAI and NT are more sensitive and measure antibodies against subtype and strain-specific antigens.
Treatment of Influenza
Neuraminidase inhibitors (first-line)
Oseltamivir (Tamiflu) and zanamivir (Relenza) inhibit neuraminidase, the enzyme that cleaves sialic acid bonds to release newly budded virions from the infected cell surface. Without NA function, new virions remain stuck on the host cell and cannot spread to infect adjacent cells. Both drugs are active against influenza A and B.
Treatment must be started within 48 hours of symptom onset for maximum benefit — the earlier the better. After 48 hours, viral replication is already near its peak, and the benefit of antiviral therapy is significantly reduced. High-risk patients (elderly, immunocompromised, pregnant women, children under 5, patients with chronic illness) should receive treatment promptly without waiting for laboratory confirmation.
M2 ion channel inhibitors (largely obsolete)
Amantadine and rimantadine block the M2 ion channel, which is required for viral uncoating during cell entry. These drugs are only active against influenza A and have no effect on influenza B. They were once widely used but are now largely ineffective due to near-universal resistance in currently circulating H3N2 and H1N1 strains. WHO and CDC no longer recommend them for routine influenza treatment.
Supportive care and complications management
- Fever and myalgia: paracetamol or ibuprofen. Aspirin must not be given to children (Reye's syndrome risk — see Complications section)
- Secondary bacterial pneumonia: requires antibiotic therapy once the causative organism is identified
- Severe influenza/pneumonia: supportive care in an intensive care setting; consider IV oseltamivir for patients unable to take oral medication
How to Remember
HA attaches, NA releases — the two surface proteins are the two ends of the replication cycle. Hemagglutinin handles the entry step (binding sialic acid to get in); neuraminidase handles the exit step (cleaving sialic acid to get out). If you can pair each surface protein with its functional job — HA in, NA out — you immediately understand why anti-HA antibodies prevent infection (blocking entry) and why neuraminidase inhibitors are antivirals (trapping progeny virions on the cell surface before they can infect adjacent cells).
Drift = typos, Shift = chapter replacement. Antigenic drift accumulates individual letter-level errors (point mutations in HA/NA), changing the surface proteins gradually, like a word slowly becoming a different word through successive typos. Antigenic shift replaces an entire gene segment at once, like substituting a whole chapter, producing a surface protein the immune system has never seen before. Epidemics come from drift; pandemics come from shift.
The pig is the mixing bowl because it speaks both languages. Pigs express both human-type (α-2,6) and bird-type (α-2,3) sialic acid receptors, so they can be infected by both human and avian influenza strains simultaneously. That dual infection in the same cell is what allows the two strains' RNA segments to mix and produce a reassortant with new surface proteins. The 2009 H1N1 pandemic strain contained gene segments from human, avian, and swine influenza lineages.
Eight segments for A and B, seven for C — the missing segment in C is neuraminidase. Influenza C has seven RNA segments because it lacks a neuraminidase gene entirely. Instead it has a single HEF (haemagglutinin-esterase-fusion) protein that performs both the HA and NA functions combined. This structural difference is also why influenza C doesn't undergo the same kind of reassortment-based antigenic shift as influenza A, contributing to its clinical mildness and the absence of influenza C pandemics.
Key Exam Facts Table
| Feature | Detail |
|---|---|
| Family | Orthomyxoviridae |
| Types | A, B, C (and D, primarily cattle) |
| Genome | Negative-sense ssRNA; 8 segments (A, B); 7 segments (C — no NA gene) |
| Envelope | Present |
| Surface glycoproteins | HA (hemagglutinin) and NA (neuraminidase) |
| HA subtypes | 18 (H1–H18) |
| NA subtypes | 11 (N1–N11) |
| Epidemics | Influenza A and B |
| Pandemics | Influenza A only (segmented genome enables reassortment across species) |
| Antigenic drift mechanism | RNA polymerase errors accumulate point mutations in HA/NA (no proofreading) |
| Antigenic shift mechanism | Reassortment of entire gene segments between two co-infecting strains |
| Pig's role in pandemic emergence | Dual α-2,6 (human) and α-2,3 (avian) sialic acid receptors allow co-infection and reassortment |
| Incubation period | 1–4 days (average 2 days) |
| Peak viral shedding | Within 24–72 hours of symptom onset |
| Onset pattern | Abrupt (vs. gradual onset in common cold) |
| Most common serious complication | Secondary bacterial pneumonia |
| Aspirin contraindication | Children with influenza — Reye's syndrome risk |
| Cell line for culture | Madin-Darby Canine Kidney (MDCK) cells |
| First-line antiviral | Oseltamivir (Tamiflu) or zanamivir (Relenza) — neuraminidase inhibitors |
| Antiviral window | Must start within 48 hours of symptom onset |
| M2 inhibitors (amantadine) | Now largely obsolete due to widespread resistance |
Where Students Get Confused
"Antigenic drift and antigenic shift are just different names for the same thing." They are completely different processes. Drift is slow, continuous, and produces annual epidemic strains through accumulated mutations. Shift is sudden, rare, and produces pandemic strains through wholesale exchange of gene segments. Drift affects influenza A and B; shift is an influenza A-only phenomenon because it requires cross-species co-infection and reassortment.
"Influenza B can also cause pandemics." It cannot. Influenza B is almost exclusively a human pathogen with a much narrower host range than influenza A. Without the ability to co-circulate widely in animal reservoirs, influenza B has limited opportunity for the cross-species reassortment that generates pandemic strains. Influenza B causes significant seasonal epidemics but has never caused a documented pandemic.
"A new flu vaccine gives you complete protection for the season." Not quite. Annual flu vaccines are designed to match the strains predicted to circulate that season, based on WHO surveillance. Vaccine effectiveness varies year to year depending on how closely the vaccine strains match the actual circulating strains after antigenic drift has occurred between the time of vaccine formulation and the start of flu season — sometimes as high as 60–70%, sometimes lower. Vaccination still substantially reduces the risk of severe disease, hospitalisation, and death even in lower-efficacy seasons.
"Tamiflu/oseltamivir works on all respiratory viruses." It is specific to influenza. Neuraminidase inhibitors target influenza NA, which is not present in other respiratory viruses like RSV, rhinovirus, SARS-CoV-2, or adenovirus. Starting oseltamivir without confirming influenza diagnosis (or in a high-index-of-suspicion clinical context) risks treating a non-influenza infection with an ineffective drug.
"Starting Tamiflu on day 3 or 4 of influenza symptoms is still useful." For most patients, the benefit diminishes substantially after 48 hours of symptom onset. Beyond 48 hours, antivirals are generally only recommended for high-risk patients who are still symptomatic and at risk of complications.
References and Further Reading
- Hutchinson, E. C. (2018). Influenza virus. Trends in Microbiology, 26(9), 809–810. https://doi.org/10.1016/j.tim.2018.05.013
- Boktor, S. W., & Hafner, J. W. (2023). Influenza. In StatPearls. StatPearls Publishing. https://www.ncbi.nlm.nih.gov/books/NBK459363/
- Pleschka, S. (2013). Overview of influenza viruses. Current Topics in Microbiology and Immunology, 370, 1–20. https://doi.org/10.1007/82_2012_272
- Dangi, T., & Jain, A. (2012). Influenza virus: A brief overview. Proceedings of the National Academy of Sciences, India. Section B, 82(1), 111–121. https://doi.org/10.1007/s40011-011-0009-6
- Paules, C., & Subbarao, K. (2017). Influenza. The Lancet, 390(10095), 697–708. https://doi.org/10.1016/S0140-6736(17)30129-0
Frequently Asked Questions
Why does the flu vaccine need to be updated every year when most other vaccines don't?
What is the difference between antigenic drift and antigenic shift?
Why do pigs play a special role in generating pandemic influenza strains?
Why must oseltamivir be started within 48 hours of flu symptoms?

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.