Nutritional Types of Bacteria
Why nearly every human pathogen falls into just one category on this classification, the discovery that revealed bacteria could "eat" rocks instead of food, and what it actually explains about how culture media are designed.
The bacteria that didn't need food at all
In the 1880s, Sergei Winogradsky was studying sulfur bacteria under the microscope and noticed something 19th-century biology had no framework for. These organisms were clearly alive and growing, with no organic food source anywhere in sight. At the time, science assumed all life needed organic carbon, either by consuming other organisms or absorbing organic molecules from the environment; the assumption was baked so deeply into biology that the very word "trophic" carried it.
Winogradsky showed something stranger was happening. The bacteria were oxidizing hydrogen sulfide, a foul-smelling inorganic gas, purely to generate energy, and using that energy to build their own bodies directly from carbon dioxide. No organic food was involved at any stage. He called this chemolithotrophy, energy from chemicals, quite literally, from rock. It was one of the first clear demonstrations that "food" for a microorganism could mean something radically different than food for anything visible to the eye.
That discovery turns out to matter for a reason far more practical than history. Once bacteria are sorted by exactly what they need for energy and carbon, a genuinely useful clinical pattern falls out immediately: virtually every organism that causes human disease lands in a single category on this list, and that single fact is the entire reason a routine bacteriology culture plate looks the way it does.
All organisms require an energy source to drive energy-consuming life processes. Energy can be tapped from light or from chemicals (organic or inorganic). Organisms that harvest radiant energy (sunlight) are called phototrophs; organisms that harvest energy from chemicals are called chemotrophs.
Separately, all organisms need a carbon source to build cell components. Organisms that use carbon dioxide as their major or sole carbon source are autotrophs; organisms that require organic compounds as their carbon source are heterotrophs. Energy source and carbon source are two independent axes, and combining them is what actually defines each nutritional type below.
Chemotrophs
Chemoorganotrophs conserve energy from organic chemicals. Thousands of different organic compounds, nearly all natural ones and most synthetic ones, can be metabolized by one microorganism or another; energy is conserved from oxidizing the compound and stored in ATP. Aerobes obtain this energy in the presence of oxygen, anaerobes in its absence, and facultative anaerobes can do either.
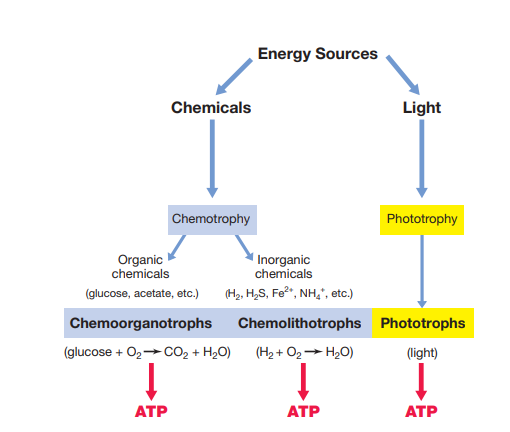 Chemolithotrophs conserve energy by oxidizing inorganic compounds instead, the phenomenon Winogradsky discovered in the hook above. Chemolithotrophy occurs only in prokaryotes and is widely distributed among Bacteria and Archaea. Common inorganic energy sources include H₂, H₂S (hydrogen sulfide), NH₃ (ammonia), and Fe²⁺ (ferrous iron), giving rise to specialized groups like the "sulfur" bacteria and "iron" bacteria. Like phototrophs, chemolithotrophs are typically autotrophs.
Chemolithotrophs conserve energy by oxidizing inorganic compounds instead, the phenomenon Winogradsky discovered in the hook above. Chemolithotrophy occurs only in prokaryotes and is widely distributed among Bacteria and Archaea. Common inorganic energy sources include H₂, H₂S (hydrogen sulfide), NH₃ (ammonia), and Fe²⁺ (ferrous iron), giving rise to specialized groups like the "sulfur" bacteria and "iron" bacteria. Like phototrophs, chemolithotrophs are typically autotrophs.
This is a genuinely good metabolic strategy: it avoids competing with chemoorganotrophs for organic energy sources, and many of the inorganic compounds chemolithotrophs oxidize (H₂, H₂S) are actually waste products excreted by chemoorganotrophs. The two groups often end up living in close association, each exploiting a resource the other can't use.
Phototrophs
Phototrophic microorganisms harvest energy from sunlight using light-absorbing pigments, which is also why their cells appear colored. Unlike chemotrophs, phototrophs don't require chemicals as an energy source at all.
Purple bacteria appeared on Earth long before oxygenic phototrophs evolved. Green sulfur bacteria were among the very first phototrophs to evolve.
Two forms of phototrophy occur in prokaryotes: oxygenic photosynthesis (produces O₂; characteristic of cyanobacteria) and anoxygenic photosynthesis (produces no O₂; characteristic of purple sulfur bacteria, green bacteria, and heliobacteria).
Within phototrophs, photolithotrophs use inorganic compounds as their electron source (e.g., Chromatium okenii), while photoorganotrophs use organic compounds like fatty acids and alcohols as electron donors instead (e.g., Rhodospirillum rubrum).
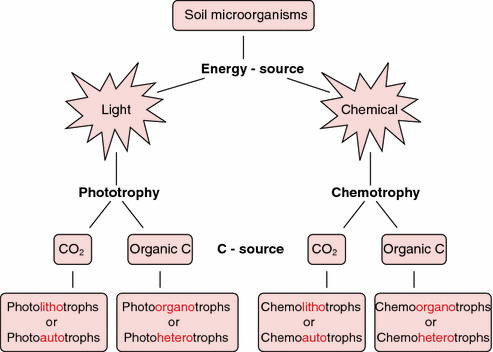 ### Putting Energy Source and Carbon Source Together
### Putting Energy Source and Carbon Source Together
| Energy source | Carbon source | Category | Example |
|---|---|---|---|
| Light | CO₂ | Photoautotroph | Cyanobacteria, Chromatium okenii |
| Light | Organic compounds | Photoheterotroph (photoorganotroph) | Rhodospirillum rubrum |
| Inorganic chemicals | CO₂ | Chemolithoautotroph | Nitrosomonas (ammonia → nitrite) |
| Organic chemicals | Organic compounds | Chemoorganoheterotroph | Virtually all human pathogens, E. coli |
Autotrophs and Heterotrophs in Practice
Chemoorganotrophs are, by definition, always heterotrophs. Most chemolithotrophs and phototrophs, by contrast, are autotrophs; Nitrosomonas, for example, oxidizes ammonia to nitrite for energy and uses that energy to fix CO₂ into cell components.
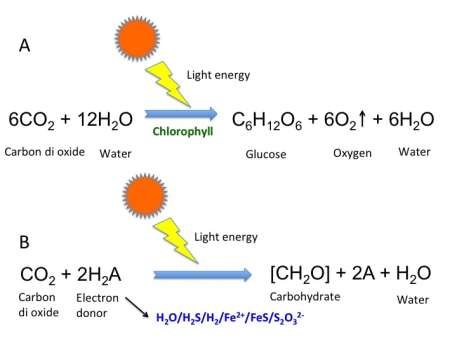 Figure: A. Oxygenic photosynthesis B. Anoxygenic photosynthesis of autotrophs (Image source: Gour Gopal Satpati)
Figure: A. Oxygenic photosynthesis B. Anoxygenic photosynthesis of autotrophs (Image source: Gour Gopal Satpati)
Autotrophs are often called primary producers, since they synthesize new organic matter from CO₂ for both their own benefit and, ultimately, everyone else's. Virtually all organic matter on Earth has been built by primary producers, particularly phototrophs, and autotrophs drive the biological cycling of elements in nature.
Heterotrophs, by contrast, rely on that organic matter, either consuming primary producers directly or living off what they excrete. All organisms that cause disease in humans, animals, and plants are heterotrophs, and they make up the majority of the microbial population in most immediate environments. Heterotrophs vary considerably in nutritional complexity: E. coli has comparatively simple requirements, while lactobacilli need a much richer, more specific set of nutrients.
Why This Classification Actually Matters
Put the two facts above together, and a specific, useful pattern emerges: nearly every organism that causes human disease is a chemoorganotrophic heterotroph. It needs organic chemicals for both energy and carbon, no light, no inorganic chemistry required. This is exactly why standard bacteriology culture media, nutrient agar, blood agar, and similar formulations, are built around organic carbon and energy sources like peptones and blood, rather than needing to supply light or a specific inorganic chemical. The classification isn't just an abstract taxonomy; it's the working logic behind how a diagnostic microbiology lab decides what to put in a culture plate in the first place.
Obligate Parasites
A small group of bacteria have nutritional and physical requirements that still aren't fully understood, and as a result, can't be cultivated on artificial media at all; they're propagated only through animal inoculation. Mycobacterium leprae, for example, can be grown by infecting the footpads of mice or, notably, nine-banded armadillos. Rickettsiae and chlamydiae are also obligate intracellular bacteria in this sense.
Spirochetes as a group are not all obligate intracellular organisms, this is a common overgeneralization worth correcting. Leptospira and Borrelia species can be cultured in vitro, though often only on specialized, fastidious media. Treponema pallidum, the cause of syphilis, is the specific spirochete that genuinely cannot be cultured on artificial media and requires animal inoculation, not the entire spirochete group.
How to Remember
The "two questions" shortcut for the whole classification. Every nutritional type on this page can be derived by answering just two independent questions: where does the energy come from (light or chemicals), and where does the carbon come from (CO₂ or organic compounds)? Answer both, and the organism's place on the entire chart follows automatically, no need to memorize six terms as unrelated vocabulary.
Anchor for the hook's payoff. Next time a culture plate looks like it's full of blood and protein rather than a sunlit fish tank, that's not incidental. Almost everything that makes people sick needs organic chemicals for both energy and carbon, exactly the chemoorganotrophic heterotroph box, and culture media are built specifically to feed that box.
Key exam facts in one table
| Fact | Detail |
|---|---|
| Two independent classification axes | Energy source (light vs. chemicals) and carbon source (CO₂ vs. organic compounds) |
| Chemoorganotroph | Energy from organic chemicals; always a heterotroph |
| Chemolithotroph | Energy from inorganic chemicals (H₂, H₂S, NH₃, Fe²⁺); typically an autotroph |
| Discovered by | Winogradsky, studying sulfur bacteria in the 1880s |
| Photoautotroph / photolithotroph | Light energy + inorganic electron donor; e.g., cyanobacteria |
| Photoheterotroph / photoorganotroph | Light energy + organic electron donor; e.g., Rhodospirillum rubrum |
| Key clinical pattern | Virtually all human pathogens are chemoorganotrophic heterotrophs |
| Why this matters practically | Standard culture media (nutrient agar, blood agar) are built around organic carbon/energy sources for exactly this reason |
| True obligate intracellular examples | Mycobacterium leprae, rickettsiae, chlamydiae, and specifically Treponema pallidum among spirochetes |
| Common overgeneralization | Not all spirochetes are obligate intracellular; Leptospira and Borrelia can be cultured, just fastidiously |
Where Students Get Confused
- Assuming all spirochetes are uncultivable. Only Treponema pallidum is a genuine obligate intracellular organism among the spirochetes; Leptospira and Borrelia can be grown in vitro on specialized media.
- Treating "autotroph" and "phototroph" as synonyms. Most phototrophs are autotrophs, but so are most chemolithotrophs, despite getting their energy from chemicals rather than light. Energy source and carbon source are separate questions.
- Assuming all chemotrophs are heterotrophs. Chemoorganotrophs always are, but chemolithotrophs are typically autotrophs, even though both are technically "chemotrophs."
- Confusing a genuinely uncultivable organism with a merely fastidious one. An obligate intracellular organism like M. leprae may lack the biosynthetic machinery to survive outside a host cell entirely; a fastidious organism just needs a richer growth medium, a fundamentally different situation.
References
- Madigan, M. T., Bender, K. S., Buckley, D. H., Sattley, W. M., & Stahl, D. A. (2018). Brock Biology of Microorganisms (15th ed.). Pearson.
- Pelczar, M. J., Chan, E. C. S., & Krieg, N. R. (2001). Microbiology: Concepts and Applications. McGraw-Hill.
- Winogradsky, S. (1887). Über Schwefelbacterien. Botanische Zeitung, 45, 489–610.
Frequently Asked Questions
What are the main nutritional types of bacteria?
What is chemolithotrophy, and who discovered it?
Why does it matter that most pathogens are chemoorganotrophic heterotrophs?
Are all spirochetes impossible to culture in a lab?
What is the difference between an autotroph and a heterotroph?
Are all chemotrophs heterotrophs?

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.