Baltimore Classification: Why Genome Type Predicts How a Virus Behaves
How knowing a virus's genome type alone predicts its mutation rate, drug vulnerabilities, and replication strategy.
Why do influenza and HIV demand a new vaccine or constant drug-resistance monitoring almost every year, while a single childhood course of the hepatitis B or HPV vaccine provides protection for decades? And why do certain antiviral drugs (like the reverse transcriptase inhibitors used for HIV) work brilliantly against some viruses but do absolutely nothing against others, no matter the dose? Both answers trace back to a single property: what kind of genome the virus carries, and how it has to convert that genome into mRNA.
This is exactly what the Baltimore system captures, and why it has remained useful for more than 50 years despite being, on the surface, "just" a classification scheme. RNA viruses that replicate using RNA-dependent RNA polymerase (an enzyme that, unlike host DNA polymerase, has no proofreading ability) make far more copying errors every replication cycle. More errors means faster mutation, which is exactly why influenza and HIV evolve quickly enough to escape last year's vaccine or this year's drug. DNA viruses, by contrast, often borrow the host's own high-fidelity DNA polymerase, inheriting its proofreading and producing a much more genetically stable virus, which is part of why a hepatitis B or HPV vaccine can protect for years without needing an annual update.
And reverse transcriptase inhibitors only work against the handful of Baltimore classes that actually use a reverse transcriptase enzyme in their replication cycle (Classes VI and VII below). Knowing a virus's Baltimore class isn't trivia, it tells you in advance what kind of vaccine durability to expect and which entire categories of antiviral drugs could plausibly work against it, before you know anything else about that specific virus.
The Baltimore system of virus classification devised by virologist & Nobel laureate David Baltimore is based on the genomic nature of the viruses. The central theme of the Baltimore system of virus classification is that all viruses must synthesize positive-strand mRNAs from their genomes toproduce proteins and replicate themselves. The precise mechanisms whereby this is achieved differ for each virus family.
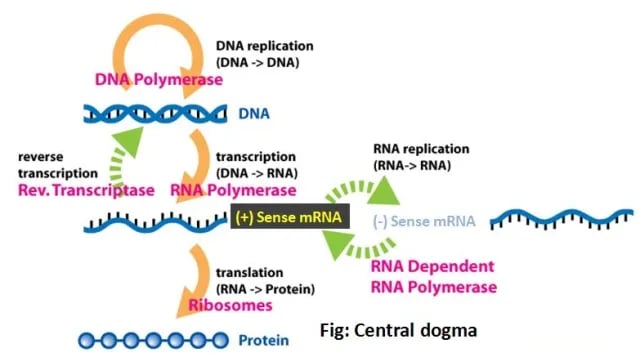 Figure: Central Dogma
Figure: Central Dogma
Why + mRNA is placed at the center?
Baltimore’s classification system is based on the central role of the translational machinery, places mRNA in the center, and describes the pathways to form mRNA from DNA or RNA genomes. Viruses can replicate DNA and/or RNA, synthesize RNA from DNA, or vice versa, but lack a complete system to make proteins, for which they have to rely on host cell ribosomes. Host cells, on the other hand, can synthesize proteins only from +mRNA strands. Irrespective of the genomic nature of viruses, all viruses must synthesize viral + mRNAs to produce viral proteins”no exception to date.”
How do you designate positive (+) and/or negative (−) strands?
The mRNA, which can serve as the template for protein synthesis, is defined as a positive (+) strand {by convention (nothing to do with charge/electricity)}. A strand of DNA of the equivalent polarity is also called the (+) strand. RNA and DNA strands complementary to the (+) strands are called negative (−) strands.
Viral genomes can be broken down into seven fundamentally different groups, which require different basic strategies for their replication depending on the nature of their genome. When originally conceived, the Baltimore scheme encompassed six classes of the viral genome. Subsequently, the gapped DNA genome of hepadnaviruses (e.g., hepatitis B virus) was discovered and incorporated into a new scheme.
Viruses can be classified into seven (arbitrary) groups:
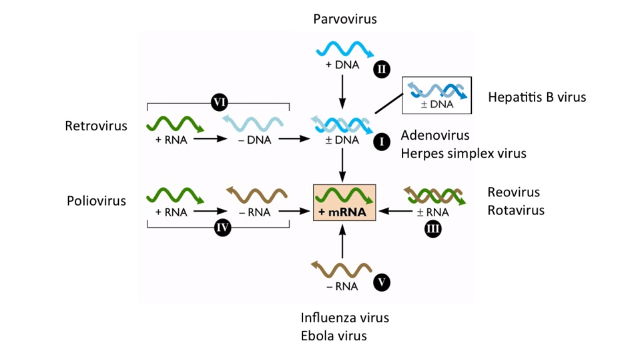 Figure: Baltimore system of classification of viruses
Figure: Baltimore system of classification of viruses
How to Remember Seven Classes
The seven classes are really just three questions, asked in sequence. Instead of memorizing seven isolated boxes, ask: (1) Is the genome DNA or RNA? (2) Is it single- or double-stranded? (3) If single-stranded RNA, is it positive-sense (already readable as mRNA) or negative-sense (needs to be flipped first)? Answering these three questions in order walks you to the correct class almost every time, classes I-V fall out directly from this logic, with VI and VII as the two "genome converts to a different nucleic acid type entirely" exceptions.
Classes VI and VII both use reverse transcriptase, but at opposite ends of the replication cycle, this is the single most commonly confused pair. Retroviruses (Class VI, like HIV) carry RNA and only make DNA after infecting a new cell, using their own reverse transcriptase to write a DNA copy (the provirus) that then integrates into the host genome. Hepadnaviruses (Class VII, like hepatitis B) do the reverse: they already carry a (gapped) DNA genome, and the RNA-to-DNA reverse transcription step happens later, during virion maturation, before the virus is even released to infect a new cell. A useful anchor: Class VI converts RNA-to-DNA after entry; Class VII's DNA genome gets completed and partially reverse-transcribed before exit. Same enzyme, opposite point in the journey.
Positive-sense RNA genomes (Class IV) are immediately infectious on their own; negative-sense genomes (Class V) are not. Because a positive-sense RNA genome already reads exactly like mRNA, host ribosomes can translate it directly the moment it enters a cell, which is why purified Class IV genomic RNA alone (with no protein) can actually start an infection. A negative-sense genome (Class V) is the mirror image of mRNA, completely unreadable by host ribosomes until the virus's own RNA-dependent RNA polymerase, which it must carry inside the virion itself, flips it into a readable positive-sense copy first. This is also why Class V viruses, unlike Class IV, must package their polymerase enzyme inside every infectious particle, without it, the genome alone is useless cargo.
I.Double-stranded DNA viruses
Some replicate in the nucleus e.g., adenoviruses using cellular proteins. Poxviruses replicate in the cytoplasm and make their own enzymes for nucleic acid replication. e.g., Adenoviruses; Herpesviruses; Poxviruses, etc
II. Single-stranded (+) sense DNA viruses
Replication occurs in the nucleus, involving the formation of a (-) sense strand, which serves as a template for (+) strand RNA and DNA synthesis. e.g., Parvoviruses
III. Double-stranded RNA viruses
These viruses have segmented genomes. Each genome segment is transcribed separately to produce monocistronic mRNAs. e.g., Reoviruses
IV. Single-stranded (+) sense RNA viruses (Picornaviruses; Togaviruses, etc)
- Polycistronic mRNA: Genome RNA = mRNA. Since the RNA is in the same sense as mRNA, the RNA alone is infectious, with no virion particle associated with polymerase. Translation results in forming a polyprotein product, which is cleaved to form the mature proteins. e.g. Picornaviruses (poliovirus, rhinovirus); Hepatitis A virus
- Complex Transcription: Two or more rounds of translation are necessary to produce the genomic RNA. e.g. Picornaviruses; Hepatitis A.
V. Single-stranded (-) sense RNA viruses
The virion RNA is a negative sense (complementary to mRNA) and must be copied into the complementary plus-sense mRNA to make proteins. This group of viruses must code for RNA-dependent RNA-polymerase and carry it in the virion to make mRNAs upon infecting the cell. e.g. Orthomyxoviruses, Rhabdoviruses, etc
- Segmented e.g. Orthomyxoviruses. The first step in replication is a transcription of the (-) sense RNA genome by the virion RNA-dependent RNA polymerase to produce monocistronic mRNAs, which also serve as the template for genome replication.
- Non-segmented e.g. Rhabdoviruses. Replication occurs as above, and monocistronic mRNAs are produced.
VI. Single-stranded (+) sense RNA viruses with DNA intermediate in life-cycle
RNA genome is (+) sense but unique among viruses in that it is DIPLOID and does not serve as mRNA but as a template for reverse transcription. e.g., Retroviruses.
Retroviruses, therefore, encode an RNA-dependent DNA polymerase (reverse transcriptase) to make the DNA provirus which then is transcribed to genomic RNA by a host enzyme, RNA polymerase II.
VII. Double-stranded DNA viruses with RNA intermediate
This group of viruses also relies on reverse transcription, but unlike the Retroviruses, this occurs inside the virus particle on maturation. On infection of a new cell, the first event is a repair of the gapped genome, followed by a transcription. e.g., Hepadnaviruses
Uses
- The elegance of the Baltimore system is that knowing only the nature of the viral genome lets you deduce the basic steps required to produce mRNA, without yet knowing anything else about that specific virus.
- It predicts a virus's likely mutation rate: RNA viruses replicating via error-prone, non-proofreading polymerases (Classes III, IV, V) tend to mutate faster than DNA viruses that borrow the host's higher-fidelity DNA polymerase (Class I).
- It narrows which categories of antiviral drugs could plausibly work: reverse transcriptase inhibitors are only relevant to Classes VI and VII; protease inhibitors target the polyprotein-cleavage strategy common in Class IV viruses.
Where Students Get Confused
Class IV and Class V genomes look similar on paper (both single-stranded RNA) but behave completely differently the moment they enter a cell. The deciding question is always: can host ribosomes read this genome directly as mRNA? If yes, it's Class IV (positive-sense, immediately infectious on its own). If no, it's Class V (negative-sense, useless to ribosomes until the virus's own polymerase converts it), and critically, the virus must carry that polymerase inside the virion, since a Class V genome alone cannot bootstrap its own conversion.
Class VI and Class VII both involve reverse transcriptase, leading students to treat them as the same thing. They're not, the order of operations is reversed (RNA-to-DNA after entry for retroviruses, versus DNA-genome-completion with reverse transcription during maturation for hepadnaviruses, see the How to Remember section above). An exam question describing "a DNA virus that uses reverse transcriptase" is testing Class VII (hepadnaviruses like HBV) specifically, not retroviruses, since retroviruses start as RNA, not DNA.
"Polycistronic" and "complex transcription" under Class IV sound like two unrelated extra facts, but they describe two different viral strategies for getting multiple proteins out of one genome. Polycistronic genomes (like poliovirus) translate the entire genome into one long polyprotein that's then cleaved into individual proteins after translation. Complex transcription describes viruses (also within Class IV) requiring multiple rounds of translation before the genomic RNA itself can be produced. Both solve the same basic problem, producing several distinct proteins from a single RNA strand, just via different mechanisms.
Key Exam Facts Table
| Class | Genome | Key replication feature | Example viruses | Clinically relevant drug target |
|---|---|---|---|---|
| I | dsDNA | Often uses host DNA polymerase; nucleus (e.g., adenovirus) or cytoplasm (poxvirus, with own enzymes) | Adenoviruses, Herpesviruses, Poxviruses | DNA polymerase inhibitors (e.g., acyclovir for herpesviruses) |
| II | ssDNA (+) | Forms (-) strand first as template for replication | Parvoviruses | Limited specific antivirals |
| III | dsRNA | Segmented genome; each segment transcribed separately | Reoviruses | Limited specific antivirals |
| IV | ssRNA (+) | Genome = mRNA directly; genome alone is infectious | Picornaviruses, Hepatitis A virus, Togaviruses | Protease inhibitors (cleave the polyprotein) |
| V | ssRNA (−) | Genome must be transcribed by virion-packaged RNA polymerase before translation | Orthomyxoviruses (influenza), Rhabdoviruses (rabies) | Polymerase inhibitors (e.g., neuraminidase inhibitors for influenza target a different step but exploit genome biology) |
| VI | ssRNA (+) with DNA intermediate | Reverse transcription occurs after cell entry; integrates as DNA provirus | Retroviruses (HIV) | Reverse transcriptase inhibitors, integrase inhibitors |
| VII | dsDNA with RNA intermediate | Gapped genome repaired, reverse transcription occurs during virion maturation | Hepadnaviruses (Hepatitis B) | Reverse transcriptase inhibitors (e.g., tenofovir) |
For a deeper explanation of the Baltimore scheme directly from virologist Vincent Racaniello, see the linked article in Further Readings below.
Further Readings
- Baltimore, D. (1971). Expression of animal virus genomes. Bacteriological Reviews, 35(3), 235–241. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC378387/
- Racaniello, V. (2009). Simplifying virus classification: The Baltimore system. Virology Blog. https://virology.ws/2009/08/12/simplifying-virus-classification-the-baltimore-system/
Frequently Asked Questions
Why do flu vaccines need to be updated every year, but the hepatitis B vaccine doesn't?
What's the difference between Class VI and Class VII viruses if both use reverse transcriptase?
Why can a purified Class IV viral genome alone start an infection, but a Class V genome can't?
Does knowing a virus's Baltimore class help predict which antiviral drugs might work against it?

Tankeshwar Acharya, MSc (Medical Microbiology)
Tankeshwar Acharya is an Assistant Professor in the Department of Microbiology at Patan Academy of Health Sciences (PAHS), Nepal, where he has been teaching and practicing clinical microbiology for over 14 years. He is the founder of Microbe Online, one of the leading free microbiology education resources on the web, covering bacteriology, mycology, parasitology, immunology, and clinical laboratory diagnostics written from direct experience in both the classroom and the diagnostic laboratory.